

Dinornis robustus (© N. Tamura)
Die Moas (Einzahl: der Moa) (Dinornithiformes) waren flugunfähige, heute ausgestorbene Vertreter der Laufvögel. In historischer Zeit waren sie mit neun Arten über beide Inseln Neuseelands verbreitet.
Als Urkiefervögel (Palaeognathae) besaßen Moas einen palaeognathen Gaumen, das heißt, sie zeigen im Gegensatz zu den so genannten Neukiefervögeln einen Pterygoid-Palatinum-Komplex (PPC), der aus den Schädelknochen Flügelbein (Pterygoid), Gaumenbein (Palatinum) und Pflugscharbein (Vomer) besteht. Sie glichen anderen Laufvögeln in einer Anzahl anatomischer Merkmale: Ihre Ober- und Unterkiefer endeten jeweils in einem dreiteiligen Hornschnabel (Rhamphotheca), deren mittlerer Kamm flach und von den jeweils seitlichen Schnabelteilen durch Furchen getrennt war. Der Nasalfortsatz des Zwischenkieferbeins war unpaarig und nicht mit dem Nasale verschmolzen. Im Beckengürtel war das Fenster zwischen Darmbein und Sitzbein (Foramen ilioischiadicum) länglich und nach hinten nicht abgeschlossen. Das breite und flache Brustbein war ohne Anzeichen eines Brustbeinkiels (Carina sterni).
Durch folgende abgeleitete Merkmale des Skeletts, in denen sich Moas von anderen Laufvögeln unterschieden, ist die Ordnung der Moas (Dinornithiformes) definiert: Flügel fehlten völlig. Vom Schultergürtel lag nur ein verkümmerter Scapulocoracoid ohne Anzeichen eines Schultergelenksockels vor. Das Becken war breit im Bereich der Hüftgelenkpfanne und dahinter. Das Brustbein zeigte gut ausgeprägte seitliche Fortsätze. Das fußwärtige Ende des Schienbeins war mit einem Sehnenkanal ausgestattet, der von einer knöchernen Verbindung überbrückt wurde. Das Laufbein besaß zwei Hypotarsalkämme anstatt eines. Moas besaßen 21 bis 23 Halswirbel, 6 Brustwirbel, 18 Beckenwirbel und 11 Schwanzwirbel, wobei die letzten Schwanzwirbel nicht zu einem Pygostyl verschmolzen waren. Eine Anzahl Schädelmerkmale kommt zu den genannten Charakteristika hinzu.
Die meisten Moa-Arten waren kurzbeinig und so groß wie ein Truthahn. Hingegen waren die Weibchen der beiden Arten der Gattung Dinornis die größten Vögel Neuseelands. Ihr Gewicht betrug etwa 180 Kilogramm, nach anderen Schätzungen bis zu 270 Kilogramm. Sie hielten den Kopf nach vorne gestreckt und in Höhe des Rückens oder darunter; dies war der Form der Wirbelsäule geschuldet, die vor den Brustwirbeln abwärts gebogen war, mit dem tiefsten Punkt zwischen den Wirbeln 12 und 16 (vom Schädel aus gezählt); die vorderen Halswirbel waren wiederum so kurz, dass sie kaum zur Erhöhung des Kopfes beitrugen. Somit besaßen die größten Vertreter der Moas, die in älteren Rekonstruktionen mit aufrechtem Hals gezeigt wurden und somit höher als ein Afrikanischer Strauß gewesen wären, Kopfhöhen von kaum mehr als zwei Metern.
Die größten bekannten Exemplare von Eiern der Moas waren 40 Zentimeter hoch und hatten ein Gewicht von etwa 4500 Gramm, damit entsprach ihr Inhalt dem von mehr als 80 durchschnittlichen Hühnereiern.
Bei den Moas gab es ursprünglich einige Missverständnisse hinsichtlich ihres Habitats. Man verglich sie mit heute lebenden großen Laufvögeln wie Straußen und Nandus und leitete daraus ab, dass sie Vögel des offenen Geländes gewesen sein müssten. So beschrieb der Geologe Julius von Haast, der sich als Erster mit diesen Vögeln intensiv auseinandersetzte, Moas als Vögel der Savanne und des Waldrandes, die kaum jemals in den Wald vordrangen. Bis in die 1950er Jahre blieb diese Theorie verbreitet. Dann erst zeigte die Palynologie, dass Neuseeland vor der Ankunft der Māori mit Ausnahme der subalpinen Zonen vollständig bewaldet war, die Grasländer also keineswegs eine natürliche Landschaft waren. Zudem ergab sich aus der Untersuchung von Mageninhalten, dass alle Arten Knospen, Blätter und Früchte von Waldpflanzen fraßen.
Moas lebten auf der Nord- und der Südinsel Neuseelands. Zwei Arten waren ausschließlich auf der Nordinsel, fünf nur auf der Südinsel verbreitet; die anderen zwei Arten fanden sich auf beiden Inseln. Nur von einer Dinornis-Art fand man auch spärliche Überreste auf Stewart Island.

Jagdszene mit Moas (Heinrich Harder)
Moas waren ausschließlich Pflanzenfresser. Durch Untersuchungen der Muskelmägen bei besonders gut erhaltenen Moa-Fossilien konnte man feststellen, dass Dinornis offenbar hauptsächlich Zweige abweideten, während Emeus und Euryapteryx weichere Kost wie Blätter und Früchte zu sich nahmen. Von Vertretern anderer Gattungen sind keine Mageninhalte bekannt, jedoch bis zu fünf Zentimeter große Magensteine (Gastrolithen). Für einen Anteil tierischer Nahrung gibt es keine Anhaltspunkte.
Moas legten ein bis zwei Eier pro Gelege. Man hat bisher etwa dreißig erhaltene Moa-Eier und unzählige Schalenreste gefunden. In den seltensten Fällen gelang es allerdings, die Eier einer Art zuzuordnen. In einem Fall fand man Überreste eines wahrscheinlich brütenden Moa mitsamt Ei, was die Zuordnung einfach machte. In anderen Fällen schloss man durch Vergleiche der Häufigkeit von Moa- und Eier-Fossilien in bestimmten Regionen auf Zusammengehörigkeit. Auffällig ist, dass Moa-Eier ungewöhnlich groß sind. So war das Ei eines Euryapteryx curtus, eines nur 20 Kilogramm schweren Moas, ebenso groß wie das des sehr viel größeren Emus. Die Eier der Dinornis-Weibchen, der größten aller Moas, waren mit Maßen von 24 × 18 cm und Volumina von etwa 4300 cm3 deutlich größer als ein Straußenei und etwa 90-mal so groß wie ein mittleres Hühnerei. Die Größe der Eier lässt den Schluss zu, dass die frisch geschlüpften Jungmoas weit entwickelt und in hohem Maße selbstständig waren.
Auch über die Laute, die Moas von sich gaben, konnte man durch die Untersuchung eines mumifizierten Euryapteryx Klarheit erlangen. Bei diesem bildet die Luftröhre eine 1,20 Meter lange Schleife, eine Struktur, die man ähnlich beim Trompeterschwan findet. Ein solches Organ ermöglichte dem Vogel, sehr laute und weit tragende Rufe zu erzeugen. Ob andere Moa-Gattungen vergleichbare Vorrichtungen hatten, ist im Moment noch Spekulation.
Vor der Ankunft des Menschen war der Haastadler der einzige Feind der Moas. Für ihn waren vor allem die kleinen und mittelgroßen Arten die Hauptbeute. Doch auch die riesigen Weibchen der beiden Dinornis-Arten fielen dem gigantischen Greifvogel gelegentlich zum Opfer. Dies weiß man von der Untersuchung der Überreste verschiedener Moas, die schwere Verletzungsspuren des Beckens aufweisen. Sie deuten darauf hin, dass der Adler seine Beute von hinten attackierte. Die Beckenknochen wurden von den Adlerkrallen regelrecht durchstochen.
Gegenüber allen anderen Vogelgruppen, auch anderen Ratiten, zeigten Moas als extreme K-Strategen eine lange Wachstumszeit sowie ein sehr spätes Erreichen der sexuellen Reife. Bei histologischen Untersuchungen der Rindengewebe verschiedener Beinknochen (Oberschenkelknochen, Tibiotarsus, Tarsometatarsus) konnten Turvey u. a. (2005) in vier der sechs bekannten Gattungen eine deutliche Zonierung der äußeren Knochenrinde und mehrere jahreszeitlich bedingte Wachstumspausen, so genannte LAGs (englisch: Lines of Arrested Growth, „Jahresringe“) nachweisen. Sie zeigen, dass die betreffenden Individuen erst nach mehreren Jahren diskontinuierlichen Wachstums ihre endgültige Größe erreichten.
Die Riesenmoas der Gattung Dinornis wichen von diesem Schema ab. Als Folge eines ausgeprägten Sexualdimorphismus kamen die Weibchen, früher fälschlich als eigene Art Dinornis giganteus klassifiziert, auf ein Gewicht von über 200 Kilogramm, während die Männchen etwa bis zu 85 Kilogramm schwer wurden (Bunce u. a. 2003, Huynen u. a. 2003). Um diese Körpergrößen zu erreichen, war das Wachstum der Vertreter der Gattung Dinornis offenbar gegenüber dem anderer Moa-Gattungen beschleunigt: Ihr äußeres Knochenrindengewebe (Kortikalis) ist von vielen Blutgefäßen durchzogen, zeigt kaum Zonierungen und weist nur in wenigen Fällen LAGs auf. Offenbar waren die Dinornis-Arten nach etwa drei Jahren ausgewachsen, während manche der kleineren Moas wie Euryapteryx bis zu neun Jahre dafür benötigten.
Um die Altersstruktur von Moa-Populationen aufzuklären, wurden die Moa-Knochen verschiedener Fossilfundorte systematisch untersucht (Turvey und Holdaway (2005)): Es stellte sich für den Fundort Bell Hill Vineyard Swamp (nahe Waikari, North Canterbury auf der Südinsel) heraus, dass nur gut ein Viertel (27,5 %) der dort gesammelten Dinornis-robustus-Knochen nicht ausgewachsenen Tieren angehörte, während der Rest von ausgewachsenen, mehrheitlich geschlechtsreifen Individuen stammte. Ähnlich wie es bei den rezenten Kiwis der Fall ist, waren bei manchen der ausgewachsenen Tiere die Mittelfußknochen nicht vollständig verwachsen, was darauf hindeutet, dass sie noch nicht geschlechtsreif waren. Der auffällig geringe Anteil fossil überlieferter Jungtiere könnte darauf hindeuten, dass ein sehr hoher Anteil der Nachkommen das Erwachsenenalter erreichte. Diese Interpretation ist jedoch nur dann zutreffend, wenn die vorliegende Grabgemeinschaft die Verhältnisse einer tatsächlichen Population repräsentiert.
Turvey u. a. interpretieren die langsamen Wachstumsraten und die Verzögerung der Geschlechtsreife bei Moas als Ergebnis einer Anpassung an Lebensräume, die frei von räuberischen Säugetieren waren. Die sehr geringe Reproduktionsrate war dafür verantwortlich, dass die Verfolgung durch den Menschen zum schnellen Aussterben führte, bevor andere Faktoren wie beispielsweise der Verlust von Lebensraum sich negativ hätten auswirken können.
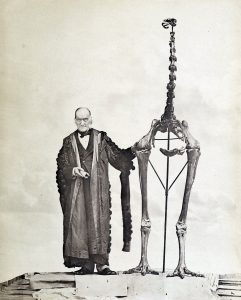
Richard Owen neben dem Skelett eines Dinornis novaezealandiae, einer der beiden Arten, deren Weibchen vormals als „Riesenmoa“ Dinornis giganteus fehlgedeutet wurden
Auffällig ist, dass Moas in den Mythen und Sagen der Māoristämme nicht vorkommen. Man konnte daher davon ausgehen, dass ihr Aussterben schon so lange zurückliegt, dass die Existenz der Riesenvögel über die Generationen in Vergessenheit geraten war.
Inzwischen lässt sich die Geschichte der Ausrottung recht gut rekonstruieren. Am Ende des 13. Jahrhunderts erreichten polynesische Einwanderer das zuvor wahrscheinlich menschenleere Neuseeland und begannen mit der Auflichtung der geschlossenen Wälder. Frühe polynesische Siedlungsplätze enthalten große Mengen von Moaknochen. Mit Ausnahme von Pachyornis australis hat man von jeder Moa-Art Überreste in Verbindung mit Menschen gefunden. Die Moas hatten abgesehen vom Haastadler keine natürlichen Feinde. Allgemein lässt sich bei Vögeln, die auf raubtierfreien Inseln leben, ein Fehlen von Flucht- oder Abwehrverhalten beobachten. So löste wahrscheinlich auch bei den Moas das Auftauchen von menschlichen Jägern weder Flucht noch Gegenwehr aus. Worthy und Holdaway mutmaßen, dass die Moa-Jagd eher einem „Einkauf im Supermarkt“ als einer Jagd gleichgekommen sein dürfte.
Schon die polynesischen Hinterlassenschaften aus der zweiten Hälfte des 14. Jahrhunderts weisen keine Moaknochen mehr auf. Dies lässt auf einen extrem kurzen Ausrottungszeitraum schließen. Die Ursprungsbevölkerung Neuseelands wird heute auf 200 Menschen geschätzt. Holdaway und Jacomb machten im Jahr 2000 den Versuch, die Ausrottung der Moas zu rekonstruieren, und kamen für manche Regionen auf extrem kurze Zeiträume; so machten sie für die Coromandel Peninsula einen Zeitabschnitt von nur fünf Jahren für die Ausrottung aller dort heimischen Moa-Spezies plausibel.
Die Ausrottung der Moas ging derart schnell vonstatten, dass die Māori nicht einmal auf die Moa-Jagd spezialisierte Waffen entwickeln mussten. Am Ende des 14. Jahrhunderts waren Moas ausgestorben. Es ist denkbar, dass einzelne Exemplare in besonders abgelegenen Regionen länger überlebten. Doch als James Cook 1769 vor Neuseeland vor Anker ging, dürften auch die letzten Moas längst verschwunden gewesen sein.
Heute gibt es manche Anhänger der Kryptozoologie, die vor allem im Fjordland nach lebenden Moas suchen. Oft gibt es auch Berichte von Wanderern, die behaupten, Moas gesehen zu haben; gelegentlich werden diese Berichte mit unscharfen Fotos untermauert. Wissenschaftler halten das Überleben der Moas bis in unsere Zeit allerdings für vollkommen ausgeschlossen.
Da die Moas aus der Überlieferung der Māori verschwunden waren, wurden sie erst anhand von Fossilfunden wiederentdeckt. Wer den ersten Knochen eines Moas fand, ist heute nicht mehr sicher nachvollziehbar. 1838 berichtete der Händler Joel Samuel Pollack von Knochenfunden, auf die ihn Māori aufmerksam gemacht hätten und aus denen er schloss, dass Emus oder Strauße einst in Neuseeland heimisch gewesen seien. Andere Reisende machten fast gleichzeitig ähnliche Entdeckungen.
In besonderer Weise widmete sich der Zoologe und Paläontologe Richard Owen (1804–1892) den Moas. Er veröffentlichte 1840 die erste Publikation über die zuvor unbekannten Großvögel (On the bone of an unknown struthious bird from New Zealand), in der er zu folgendem Urteil kam: „Ich bin willens, meine Reputation für die Folgerung aufs Spiel zu setzen, dass es in Neuseeland einst straußenartige Vögel gegeben hat oder noch gibt, die in der Größe einem heutigen Strauß nahe oder gar gleichkamen.“ Owen beschrieb die meisten der heute bekannten Moa-Arten und veröffentlichte im Laufe der folgenden fünfzig Jahre fast 50 weitere Artikel über Moas.
Weitere große Beiträge zur Moa-Forschung leistete der deutschstämmige Naturforscher Julius von Haast, der eine Sammlung von Moa-Fossilien aufbaute und neben Verdiensten bei der Beschreibung weiterer Arten über die Lebensweise der Moas spekulierte. Obwohl viele seiner Mutmaßungen heute widerlegt sind, findet man sie oft zitiert. So geht auf Haast die heute für unwahrscheinlich gehaltene Hypothese zurück, dass nicht die Māori die Moas ausrotteten, sondern ein vorher in Neuseeland lebendes Volk, das er die „Moa-Jäger“ nannte.
Das Wort „Moa“ bedeutet in vielen polynesischen Sprachen schlicht Henne. Die Anwendung dieses Namens auf die Riesenvögel geht vermutlich auf den Missionar William Colenso zurück, der nach einem Besuch bei den Māori in Waiapu von einem Mythos berichtete, an den die Einheimischen glaubten. Dieser berichte von einem riesenhaften Huhn mit dem Gesicht eines Menschen, das von zwei riesigen Echsen bewacht wurde und jeden Eindringling zu Tode trampele. Dieses Wesen würde Moa genannt. Aufgrund ähnlicher Legenden wurden anfangs auch die Māori-Wörter Tarepo und Te Kura auf die Riesenvögel bezogen. Letztlich setzte sich die Bezeichnung Moa durch.


