
Die Erforschung der spätquartären Avifauna ist eine Teildisziplin der Paläornithologie, die sich mit Vogeltaxa beschäftigt, die zwischen dem Jungpleistozän und dem Jahre 1500 (Beginn der Besiedelung Amerikas und der Welt durch die Europäer) ausgestorben sind. Ein weiterer Aspekt des Forschungsfeldes ist der Zwerg- bzw. Riesenwuchs von Vögeln, der Verlust der Flugfähigkeit und die Abstammungslinie von prähistorischen zu rezenten Taxa.
Führende Vertreter auf diesem Gebiet sind oder waren Trevor H. Worthy, Storrs Lovejoy Olson, Pierce Brodkorb, Jean-Christophe Balouet, David William Steadman, Anthony Cheke, Helen Frances James, Julian Pender Hume und Alice Cibois, deren Forschungsarbeit in den Höhlen und Sedimentschichten Australiens, der Maskarenen, Hawaiis, Neuseelands, der Karibik, St. Helenas, Ascensions, Madagaskars, der Mittelmeer-Inseln, der Kanarischen Inseln, Melanesiens und Polynesiens unschätzbare Daten über die erloschene Avifauna dieser Regionen zu Tage förderte.
Einer der Pioniere der Paläornithologie war Richard Owen, der sich als erster mit der Untersuchung von Moa-Knochen beschäftigte.
In der Zeit des späten Quartärs (ab 40.000 v. Chr. bis 1500 n. Chr.) sind unzählige Vogelarten ausgestorben, die man heute nur von Knochenfunden aus Pleistozänablagerungen oder vermischten Pleistozän/Holozänablagerungen kennt und deren Alter man nur sehr schwierig mit Hilfe der Radiokohlenstoffdatierung oder DNA-Analysen feststellen kann. Bei manchen Inselformen vom Atlantik, der Karibik, dem Mittelmeer, dem indoaustralischen und pazifischen Bereich geht man bei einigen Arten sogar davon aus, dass sie bis kurz vor dem 16. Jahrhundert überlebt haben könnten.
Nach Überzeugung vieler Wissenschaftler ist vor allem der Mensch entweder direkt oder indirekt für das Aussterben vieler Vogelarten in dieser Zeit verantwortlich. Wissenschaftlich nachgewiesen ist dies bei der ersten Besiedelung der Pazifik-Inseln vor 3500 Jahren, wo die endemische Vogelwelt Schaden durch Krankheitserreger, Überjagung, Eiersammeln, Lebensraumzerstörung oder die eingeschleppte Pazifische Ratte (Rattus exulans) nahm. Auch für Australien nehmen einige Wissenschaftler dies an, was jedoch in der Fachwelt äußerst umstritten ist
Elefantenvögel

Elefantenvogel (Avifauna, Alphen aan den Rjin)
Die Elefantenvögel (Aepyornithidae, auch Madagaskar-Strauße oder madagassisch Vorompatras) sind eine ausgestorbene Familie der Laufvögel mit den drei Gattungen Aepyornis, Mullerornis und Vorombe. Sie ist durch Fossilien und zahlreiche subfossile Eifunde von der Insel Madagaskar vor der Ostküste Afrikas bekannt. Die erstmalige Entdeckung eines Fossils dieser Vögel gelang dem Franzosen Alfred Grandidier während einer seiner Forschungsreisen auf der Insel zwischen 1865 und 1870.
Die Aepyornithiden wiesen die typische cursoriale (auf Schnelläufigkeit abgestimmte) Anatomie eines Bodenvogels auf: Die Beine waren lang und die Zehen kurz, während der Flugapparat nahezu gänzlich zurückgebildet wurde. Die Armknochen sind bis auf den Oberarmknochen verloren, das Brustbein ist flach und ungekielt. Der Hals ist lang und trägt einen verhältnismäßig kleinen Schädel. Aepyornis maximus war der größte Vertreter des Taxons und dürfte zwischen 2,7 und 3 m Höhe und über 400 kg Gewicht erreicht haben. Damit handelt es sich um einen der größten bekannten Vögel der Erdgeschichte und den größten Vogel in historischer Zeit, nur übertroffen von dem etwa gleich hohen, jedoch mutmaßlich schwereren Dromornis stirtoni aus dem Miozän oder vergleichbar mit Brontornis burmeisteri, der zwar ebenfalls etwa gleich hoch wurde, jedoch ein geringer geschätztes Gewicht von ca. 350 bis 400 kg erreichte. Die Riesenmoa (Dinornis) mit einer Scheitelhöhe von bis zu 3,6 m und einem geschätzten Gewicht von um die 250 kg, die am Ende des 14. Jahrhunderts ausgerottet wurden, waren die einzigen Vögel, die während historischer Zeit ähnliche Ausmaße erreichten. Von Aepyornis maximus sind auch subfossile Eierschalen und komplette Eier bekannt, deren Umfang in einigen Fällen bis zu einem Meter und die Länge 34 cm beträgt.
Die Aepyornithiden waren auf Madagaskar endemisch. Madagaskar trennte sich bereits in der Kreidezeit vom Kontinent Afrika, was einige auf einen Verlust der Flugfähigkeit der Vorompatra in situ schließen ließ. Phylogenomische Studien stützen die Paraphylie der Laufvögel in klassischer Auffassung und suggerieren einen unabhängigen Verlust der Flugfähigkeit innerhalb der Laufvögel. Mitochondriale DNA legt nahe, dass Elefantenvögel innerhalb der Urkiefervögel nahe mit einem Taxon der Kiwis, Emus und Kasuaren verwandt sind.
Es gibt keine direkten Belege der Nahrung, die Elefantenvögel üblicherweise zu sich nahmen. Die meisten heute lebenden Laufvögel sind Omnivoren. Es gibt jedoch dick beschalte Regenwaldfrüchte auf Madagaskar, die nach Meinung einiger auf die Verdauung durch Laufvögel ausgelegt waren. So weist etwa die Frucht der heute stark bedrohten Kokospalme Voanioala gerardii eine derartige dicke Schale auf, und einige madagassische Palmfrüchte zeigen eine dunkelblau-violette Farbe (z. B. Revenea louvelii und Satranala decussilvae), ähnlich jenen Früchten, die von Kasuaren bevorzugt werden.
Die Familie Aepyornithidae wird in drei Gattungen unterteilt:
Aepyornis, Mullerornis und Vorombe
Bisher wurden vier Arten beschrieben.
Elefantenvögels waren einst häufig auf Madagaskar und über die ganze Insel verbreitet. Man stimmt weitestgehend darin überein, dass das Aussterben des Elefantenvogels auf menschliche Einflüsse zurückzuführen ist. Womöglich dienten die großen Vögel als wichtige Fleischlieferanten. Es gibt mehrere Belege von geschlachteten Elefantenvögeln. Insbesondere die Eier der Vorompatras dürften gefährdet gewesen sein; es gibt Belege dafür, dass sie als Mahlzeiten zubereitet wurden. Wahrscheinlich spielte auch die von den Einwohnern betriebene Brandrodung eine zusätzliche Rolle, da sie große Flächen des Lebensraums des Elefantenvogels zerstörte. Des Weiteren ist möglich, dass Krankheiten, übertragen durch eingeführtes Geflügel, eine Rolle spielten.
Das Datum des Aussterbens der Elefantenvögel ist nicht sicher belegt. Archäologische Beweise belegen ein Fortleben bis mindestens zum Jahr 1000. Gelegentlich wird über ein Überleben des Vorompatra bis in das 17. Jahrhundert spekuliert. Der erste französische Gouverneur Madagaskars, Étienne de Flacourt, berichtete von einem großen Vogel, der die Eier eines Straußes legte und die Ampatres bewohnte. Dieser Vogel suche sich die entlegensten Regionen, um nicht von Menschen bedroht zu werden.
Donnervögel
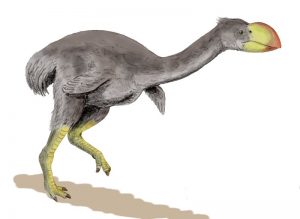
Stirton-Donnervogel (© N. Tamura)
Die Donnervögel (Dromornithidae) waren große, flugunfähige Vögel aus Australien und Tasmanien. Sie sind auch unter dem Namen Riesenemus oder in Australien als Mihirungs bekannt und gehören zu den Gänsevögeln (Anseriformes). Die ältesten Funde sind ca. 20 Millionen Jahre alt.
Dromornis australis und der Stirton-Donnervogel (Dromornis stirtoni) wurden über eine halbe Tonne schwer und 3 Meter hoch. Genyornis newtoni lebte noch im Pleistozän und war den Aborigines bekannt. Knochenfunde in menschlichen Siedlungen zeigen, dass er gejagt wurde. Er starb vor ca. 11.000 Jahren aus.
Ihr Aussterben ist auf die Klimaveränderungen, Buschbrände und möglicherweise die Jagd zurückzuführen.
Der Stirton-Donnervogel war mit einer Höhe von 2,7 bis 2,8 Metern und einem geschätzten Gewicht von bis zu 570 kg der schwerste bekannte Vogel der Erdgeschichte. Vergleichbare Werte werden Brontornis burmeisteri zugeschrieben, der zwar etwa gleich hoch wurde, jedoch lediglich ein geschätztes Gewicht von 350 bis 400 kg erreichte. Dem aus Madagaskar stammenden Aepyornis maximus schreibt man bei etwa gleicher Wuchshöhe ein erreichbares Gewicht von 450 kg zu. Schließlich ist die neuseeländische Gattung Dinornis vergleichbar, deren Weibchen aufgerichtet immerhin 3,6 Meter Scheitelhöhe, jedoch maximal 250 kg Masse erreicht haben sollen.
Sein Schädel erreichte eine Länge von 48 bis 52 Zentimeter und war etwa 25 % größer als der des zweitgrößten Donnervogels Bullockornis. Der Schnabel war, wie bei dem paläogenen, in Europa und Nordamerika lebenden Gastornis, hochgewölbt und im Vergleich zum Rest des Schädels sehr groß, aber nicht so hoch und konkav wie bei Bullockornis.
Im Unterschied zu den anderen Donnervögeln sind bei Dromornis und bei Bullockornis der erste (Atlas) und der zweite Halswirbel (Axis) zu einem einzigen Knochen zusammengewachsen. Ein unterer Halswirbel war 16,5 cm breit und 12 cm lang. Das Brustbein war breiter als lang.
Der Stirton-Donnervogel hat sich möglicherweise carnivor ernährt. Sein Lebensraum war das offene subtropische Waldland im Norden Australiens.
Nach dem Stirton-Donnervogel (Dromornis stirtoni) war Bullockornis planei die zweitgrößte Art der Gruppe, wurde zwei bis eventuell 2,8 Meter hoch und konnte ein Gewicht von wahrscheinlich 300 kg erreichen. Zeichnerische und durch reproduzierte Knochenteile anderer Donnervogel ergänzte Skelettrekonstruktionen in Museen erwecken einen Eindruck vom Aussehen der Vögel, sind aber aufgrund der unvollständigen Fossilfunde zum Teil spekulativ.
Im Unterschied zu den anderen Donnervögeln sind bei Bullockornis und Dromornis der erste (Atlas) und der zweite Halswirbel (Axis) zu einem einzigen Knochen zusammengewachsen.
Nach neueren, 1998 beschriebenen Fossilien besaß Bullockornis einen außergewöhnlich wuchtigen, mit einer ausgeprägten Muskulatur versehenen Kopf und einen Schnabel, groß genug, um einen Fußball zu verbergen. Der Schnabel war, wie bei dem paläogenen, in Europa und Nordamerika lebenden Gastornis, hochgewölbt und im Vergleich zum Rest des Schädels sehr groß. Die Augen waren klein. Kopf und Schnabel erreichten zusammen eine Länge von einem halben Meter. Der australische Paläontologe Stephen Wroe vermutet deshalb, dass Bullockornis, im Unterschied zu seinen pflanzenfressenden Verwandten, ein Fleisch- oder Aasfresser gewesen sein könnte.
Moa-Nalos
Die Moa-Nalos waren flugunfähige endemische Entenvögel (Anatidae) der Hawaii-Inseln.
Das Wort Moa-Nalo setzt sich zusammen aus Hawaiisch „Moa“ für Geflügel und „nalo“, was verloren oder vergessen heißt.
Vor der Ankunft des Menschen auf dem hawaiischen Archipel waren die Moa-Nalos die größten Pflanzenfresser der Inselgruppe und nahmen eine ähnliche ökologische Nische ein wie die verschiedenen Riesenschildkrötenarten auf den Maskarenen, den Seychellen und den Galapagosinseln. Sie waren darauf spezialisiert, Pflanzenfasern im hinteren Dickdarm zu verdauen.
Der Kauai-Moa-Nalo (Chelychelynechen quassus) hatte einen hohen, breiten Schnabel, der – ähnlich einem Schildkrötenmaul – dem Abweiden von Kräutern diente. Die längeren nach unten gebogenen, spitzen Schnäbel der anderen Moa-Nalo-Arten hatten knöcherne Pseudozähne, um Gras und Zweige fressen zu können. Als Abwehr gegen die Moa-Nalos wie auch gegen die Hawaiigänse (Nēnēs) hat zum Beispiel die zu den Lobelioideae (Lobeliengewächsen) gehörende Pflanzengattung Cyanea Stacheln sowie unterschiedlich aussehende Blätter (Heterophyllie) entwickelt.
Der Schultergürtel und die Flügel waren extrem reduziert, noch stärker als bei den Riesengänsen der Gattung Cnemiornis auf der neuseeländischen Südinsel. Auch der Brustbeinkamm fehlte völlig. Die Hinterbeine waren entsprechend stärker ausgebildet. Ihre Knochen sind durchweg sehr groß und kräftig. Schlüssel- und Rabenbein sind deutlich zurückgebildet. Bei einem Teil der Arten ist das Rabenbein entweder mit dem Schulterblatt, mit dem es einen rechten Winkel bildet, oder mit dem Brustbein verwachsen. Das Gabelbein ist sehr schwach ausgebildet und fehlt möglicherweise bei einer Population. Die Flügelknochen sind sehr klein im Vergleich zur Größe der Vögel und oft deutlich verformt. Elle und Speiche sind oft zu einem einzigen Knochen verwachsen, die Lücke zwischen den beiden Knochenteilen des Carpometacarpus ist oft geschlossen und dieser Knochen ist oft mit der ersten Phalanx des großen Fingers verwachsen. Die Schnäbel sind groß, kräftig und ungewöhnlich geformt.
Wegen der extremen Rückbildung der Flügel und des Schultergürtels ist eine systematisch korrekte Einordnung der Arten schwierig. Olson und James vermuteten 1991 aufgrund der Form der Syrinx zu der Annahme, dass es sich bei den Moa-Nalos eher um Enten (Anatinae) als um Gänse (Anserinae) handeln müsse. Livecey kam 1996 durch Untersuchung des Skeletts zu dem Ergebnis, dass sie eine Schwestergruppe der Gänse und Schwäne seien. Inzwischen wurde 1999 durch genetische Untersuchungen festgestellt, dass ihre nächsten lebenden Verwandten die Schwimmenten (Anantini) sind.
Die Moa Nalos spalteten sich vermutlich vor über drei Millionen Jahren von den übrigen Schwimmenten (Anantini) ab. Das heißt, dass sie vermutlich damals Kauaʻi und/oder Oʻahu erreichten und später eine Art, die noch nicht ganz flugunfähig war, nach Molokai auswanderte, von wo aus sie im Pleistozän zu Fuß auf die Inseln Lanai und Maui auswandern konnte, die damals mit Molokai eine zusammenhängende Insel bildeten. Hawaii konnte sie nie besiedeln, da sie bei der Entstehung dieser Insel längst völlig flugunfähig war.
Man unterscheidet vier Arten:
Der Kauai-Moa-Nalo (Chelychelynechen quassus Olson & James, 1991) lebte auf Kauai.
Der Moa-Nalo (Thambetochen chauliodous Olson & Wetmore, 1976) lebte auf Molokai, Maui und Lanai.
Der Oahu-Moa-Nalo (Thambetochen xanion Olson & James, 1991) lebte auf Oʻahu.
Der Maui-Moa-Nalo (Ptaiochen pau) lebte auf Maui.
Die Moa-Nalos wurden etwa vor 1600 Jahren als eine der ersten Vogelarten der Inselgruppe von den polynesischen Siedlern ausgerottet.


