
| ohne Rang: | Holozoa |
| ohne Rang: | Vielzellige Tiere (Metazoa) |
| Stamm: | Scheibentiere (Placozoa) |
| Gattung: | Trichoplax |
| Art: | Trichoplax adhaerens |
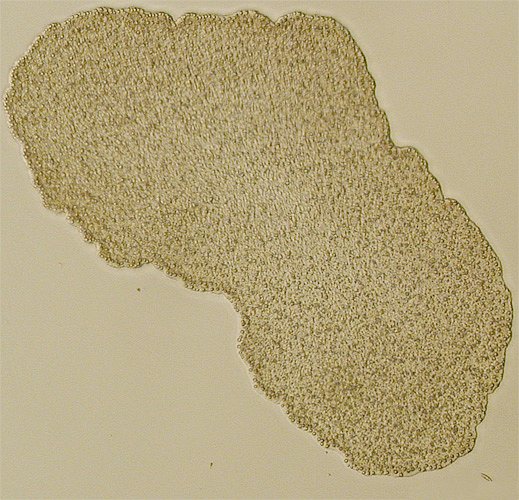
Trichoplax adhaerens (Oliver Voigt)
Trichoplax adhaerens wurde 1883 von dem deutschen Zoologen Franz Eilhard Schulze in einem Meerwasser-Aquarium des Zoologischen Instituts in Graz entdeckt. Der von ihm verliehene Gattungsname des Tiers leitet sich aus dem altgriechischen θρίξ thrix „Haar“ und πλάξ plax „Platte“ ab, das lateinische Artepithet adhaerens bedeutet „haftend“, so dass sich der Artenname etwa mit „anhaftende haarige Platte“ übersetzen lässt.
Die Placozoa oder Plattentiere sind die strukturell einfachsten aller vielzelligen Tiere (Metazoa) und bilden einen eigenen Tierstamm. Der wissenschaftliche Name bedeutet wörtlich übersetzt „flache Tiere“, im Deutschen wird gelegentlich der Name Scheibentiere verwendet. Anhand genetischer Daten gilt es als sicher, dass die Placozoa zahlreiche Arten umfassen. Da sie sich aber morphologisch nicht unterscheiden lassen, wurde bisher nur Trichoplax adhaerens als einzige Art beschrieben.
Lange Zeit hielt man die bisher nicht in ihrem natürlichen Lebensraum beobachteten Tiere für Jugendstadien von Nesseltieren (Cnidaria): Länger als ein halbes Jahrhundert wurden sie als Planula-Larven der Hydrozoen-Art Eleutheria krohni gedeutet. Erst Arbeiten aus den 1970er Jahren, unter anderem durch den Tübinger Protozoologen Karl G. Grell, klärten die frühen Phasen der Embryonalentwicklung der Tiere auf und trugen maßgeblich zur Errichtung eines eigenen Tierstamms für sie bei.
Placozoa haben, wie ihr Name andeutet, in der Regel einen stark abgeflachten, scheibenförmigen Körper. Er misst im Durchmesser in den meisten Fällen weniger als einen halben Millimeter. Vereinzelt erreichen Exemplare Größen von 2 bis 3 Millimetern – selten mehr. Die Scheibendicke beträgt meistens nur etwa 25 Mikrometer. Mit bloßem Auge sind die gräulich gefärbten, im Gegenlicht transparenten Tiere meist gerade noch zu erkennen. Oberflächlich sehen sie wie große Amöben aus und ändern wie diese beständig ihre äußere Form. Selten treten daneben kugelförmige Lebensstadien auf, die möglicherweise dem passiven Transport in neue Lebensräume dienen.
Wie die Zuordnung zu den Gewebelosen schon andeutet, fehlen Trichoplax adhaerens Gewebe und Organe; es gibt auch keine ausgeprägte Körpersymmetrie, so dass sich weder vorne oder hinten noch links oder rechts unterscheiden lassen.
Sowohl strukturell als auch funktionell lassen sich bei Trichoplax adhaerens eine Rücken- oder Dorsalseite und eine Bauch- oder Ventralseite unterscheiden. Beide bestehen aus einer einlagigen, nach außen von Schleim bedeckten Zellschicht und erinnern vor allem wegen der zwischen den Zellen bestehenden Verbindungen, der Gürteldesmosomen, an Epithelgewebe. Die Zelllagen der Placozoa besitzen aber im Gegensatz zu einem echten Epithel keine Basallamina; darunter versteht man eine aus extrazellulärem Material bestehende, dem Epithel von innen unterliegende dünne Schicht, die es versteift und vom Innenraum des Körpers abtrennt. Das Fehlen dieser ansonsten bei allen Tieren außer einigen Schwämmen zu findenden Struktur lässt sich funktionell begründen: Eine steife Trennschicht würde die amöboiden Formänderungen von Trichoplax adhaerens unmöglich machen. Um den Unterschied zu betonen, spricht man statt von einem Epithel daher manchmal bei den Placozoa von einem Epitheloid.
Ein ausgewachsenes Individuum besteht aus bis zu tausend Zellen, die sechs verschiedenen Zelltypen zugeordnet werden können (früher waren nur vier bekannt). Die einfach begeißelten Zellen des rückseitigen Epitheloids sind außen abgeflacht und bilden hier einen plattenartigen Zellverband, von dem die Zellkörper ins Innere vorragen, sie enthalten kleine Lipidkörperchen, möglicherweise Abwehrstoffe gegenüber Fressfeinden. Zwischen ihnen befinden sich wenige Zellen mit becherförmigem Zellkern, die einen kristallinen Einschluss enthalten und deshalb Kristallzellen genannt werden; sie könnten eine Funktion als Sinneszellen besitzen. Die Zellen der Bauchseite sind ebenfalls nur einfach begeißelt, sie haben eine langgestreckte säulige Form mit kleinem Querschnitt an der Oberfläche, so dass die Geißeln bauchseitig sehr eng beieinander stehen und eine bewimperte „Kriechsohle“ bilden. Ihre zahlreichen Mikrovilli deuten darauf hin, dass sie auch an der Verdauung von Beute beteiligt sind. Dazwischen, vor allem randlich auf der Ventralseite, befinden sich ebenfalls mit einem Cilium bewimperte Drüsenzellen, die Neuropeptide synthetisieren können. Diese könnten an der Steuerung und Regulation der Bewegung des Tiers beteiligt sein. Ein dritter Typ unbewimperter Zellen, die sogenannten lipophilen Zellen, erstrecken sich mit ihrem Zellkörper weit ins Innere des Organismus, sie sind in der Mitte der Kriechsohle am häufigsten, also genau entgegengesetzt wie die Drüsenzellen verteilt. Vermutlich synthetisieren diese Zellen Verdauungsenzyme.
Zwischen den beiden Zellschichten befindet sich ein flüssigkeitsgefüllter Innenraum, in dem ein lockeres Netzwerk von, überwiegend einschichtig angeordneten, Faserzellen liegt. Diese bestehen aus dem Zellkörper mit sechs oder mehr langen Fortsätzen, die sich am Ende fein verzweigen. Entgegen früheren Annahmen bilden die Faserzellen wohl kein Synzytium aus. Die Fortsätze ziehen sich zu den Zellen beider Epithelien und zu anderen Faserzellen, sie verknüpfen so alle somatischen Zellen zu einem Netzwerk. Die Kontaktstellen sind morphologisch unspezialisiert, sie bilden keine Synapsen oder ähnliche Bildungen aus. Spärlich in die Faserzellen eingestreute Scheidewände (Septen) könnten aber eine Funktion bei der elektrischen Erregungsleitung besitzen, entsprechende Funktionen sind von ähnlichen Strukturen bei anderen Tierarten bekannt. Stäbchenförmige Einschlüsse könnten endosymbiontische Bakterien unbekannter Funktion sein. Entgegen früheren Annahmen besitzen die Fortsätze keine Myofibrillen, eine Rolle, analog Muskelzellen, bei der Bewegung des Tiers ist damit eher unwahrscheinlich. Ihre Oberfläche trägt, nach Immunfluoreszenz-Beobachtungen, das Cadherin TaCDH. Es gibt Hinweise darauf, dass unterschiedliche genetische Linien der Placozoa unterschiedliche Faserzellen enthalten.
Soweit bisher bekannt, existiert keine extrazelluläre Matrix wie man sie bei Nesseltieren oder Rippenquallen als Mesogloea oder bei Schwämmen als Mesohyl findet.
Anders als bei den Gewebetieren wurden pluripotente Stammzellen, also solche, die sich in andere Zelltypen umwandeln können, noch nicht eindeutig nachgewiesen.
Alle Zellkerne der Placozoa-Zellen enthalten zwölf nur etwa zwei bis drei Mikrometer große Chromosomen. Drei Paare sind metazentrisch, die anderen acrozentrisch, das heißt, das Zentromer, die Ansatzstelle der Spindelfasern bei der Zellteilung, liegt in der Mitte beziehungsweise am äußeren Rand des jeweiligen Chromosoms. Die Zellen des Fasersynzytiums sind eventuell tetraploid, enthalten also einen vierfachen Chromosomensatz.
Insgesamt enthält ein einfacher Chromosomensatz von Trichoplax adhaerens weniger als 50 Millionen Basenpaare (98 Millionen Basenpaare des diploiden Organismus) und bildet damit das kleinste tierische Genom; die Zahl der Basenpaare des Darmbakteriums Escherichia coli ist nur um einen Faktor 10 kleiner. Es werden zwischen 11.000 und 12.000 proteinkodierende Gene abgeschätzt. Im Gegensatz zu anderen Arten mit kleinem Genom ist dasjenige von Trichoplax nicht an Introns verarmt; das ist ein Indiz gegen eine nachträgliche Verkleinerung.
Die genetische Ausstattung von Trichoplax adhaerens ist noch nicht sehr gut erforscht; einige Gene wie Brachyury oder Tbx2/3, die homolog zu entsprechenden Basenpaarsequenzen bei Gewebetieren sind, konnten allerdings bereits nachgewiesen werden. Von besonderer Bedeutung ist Trox-2, ein Placozoa-Gen, das vermutlich den bei Nesseltieren unter dem Namen Cnox-2 und bei den zweiseitig-symmetrischen Bilateria als Gsx bezeichneten Genen homolog ist, es gehört demnach zu den ParaHox-Genen. Als Homöobox- Gen, das zu den Hox-Genen gehört, spielt es in der Embryonalentwicklung bei Gewebetieren eine Rolle bei der Ausbildung und Körperdifferenzierung entlang der Symmetrieachse; bei den Nesseltieren legt es etwa die Lage von Mund- (oraler) und mundabgewandter (aboraler) Seite des Tieres fest. Placozoa besitzen allerdings, wie angesprochen, keine Symmetrieachsen. Im Trichoplax-Körper wird das Gen in einer ringförmigen, den Körper umgebenden Zone abgelesen. Vergleich der Genanordnung anderer Gene auf dem Chromosomenstrang liefert außerdem Indizien dafür, dass Trochoplax wohl ein weiteres, echtes Hox-Gen besessen haben könnte, das später verloren gegangen ist – ein Indiz für eine sekundäre Vereinfachung.
Das Trichoplax-Genom enthält darüber hinaus zahlreiche weitere Gene, die ortholog zu Genen für Transkriptionsfaktoren sind, die bei Bilateriern (wie auch dem Menschen) an der Ausbildung von Körperachsen beteiligt sind, obwohl diese bei den Placozoa nicht differenziert sind. So sind Gene für den Wnt-Signalweg und den BMP/TGF beta Signalweg vorhanden, der bei Bilateria an der Ausbildung der Körperlängsachse und der Dorsal-Ventral-Achse beteiligt ist. Obwohl spezialisierte Nervenzellen nicht ausgebildet sind, sind Proteine für Neurotransmitter und für Nervenzellen typische Ionenkanäle vorhanden. Wie typisch für Mehrzeller (aber auch schon in vielen einzelligen oder Zellkolonien bildenden Kragengeißeltierchen vorhanden) werden Gene für zahlreiche Zelladhäsionsmoleküle gefunden, darunter auch solche, die an Moleküle der extrazellulären Matrix binden, obwohl eine solche in Trichoplax noch nicht sicher nachgewiesen wurde – möglicherweise ist diese doch vorhanden, aber den bisherigen Nachweismethoden entgangen.
Genaue Verbreitungsangaben existieren für die Placozoa nicht, die Tiere wurden aber unter anderem im Roten Meer, dem Mittelmeer, der Karibik, vor Hawaii, Guam, Samoa, Japan, Vietnam oder Papua-Neuguinea und am Großen Barriere-Riff vor der australischen Ostküste nachgewiesen. Sie sind, soweit bekannt, in allen tropischen und subtropischen Meeren, nördlich bis zum Mittelmeer, bei Wassertemperaturen zwischen 10 und 32 °C weit verbreitet. Regelmäßig wird Trichoplax adhaerens auch in Meerwasser-Aquarien „gefunden“, so etwa im südenglischen Plymouth oder im US-amerikanischen Bundesstaat Florida in Miami.
Eine direkte Nachsuche im Meer ist für die sehr kleinen und unauffälligen Tiere unmöglich. Zum Nachweis existieren zwei Methoden: Entweder werden Hartsubstrate wie Steine im Lebensraum eingesammelt und später im Aquarium näher untersucht, oder es werden künstliche Substrate, meist Objektträger aus Glas, im Lebensraum exponiert, wobei man auf eine spontane Besiedlung durch die Tiere hofft. Die bisherigen Feldfunde stammen aus Flachwasser der Meeresküsten, bis in eine Tiefe von etwa 20 Meter, und Gezeitenzonen tropischer und subtropischer Meere, wo sich die Tiere auf Substraten wie Stämmen oder Wurzeln von Mangroven, Weichtier-Schalen, Bruchstücken von Steinkorallen oder einfach auf Felsbrocken finden. Sie können sowohl Brackwasser-beeinflusste Lebensräume mit einer Salinität bis 2 % wie auch etwas aufgesalzenes, konzentriertes Meerwasser mit einer Salinität bis über 5 % überleben. Ihre Häufigkeit ist in den Sommermonaten höher.[8] Die Häufigkeit des Vorkommens steigt, nach Modellberechnungen, mit steigender Temperatur und steigender Salinität an, wobei eher nährstoffarmes Wasser bevorzugt wird. Die unterschiedlichen genetischen Linien (die vermutlich verschiedenen Arten entsprechen) zeigen dabei unterschiedliche Vorlieben.
Trichoplax adhaerens ernährt sich von kleinen Algen, insbesondere Grünalgen (Chlorophyta) der Gattung Chlorella, Kryptomonaden (Cryptophyta) der Gattungen Cryptomonas oder Rhodomonas, Cyanobakterien (Cyanobacteria) wie Phormidium inundatum, aber auch von abgestorbenen Teilen anderer Organismen. Dazu bilden sich auf der Bauchseite um die Nahrungspartikel eine oder mehrere kleine Taschen aus, in die von den Drüsenzellen Verdauungsenzyme abgegeben werden; die Tiere entwickeln also gewissermaßen einen vorübergehenden „Außenmagen“. Die aufgeschlossenen Nährstoffe werden dann durch Pinocytose („Zell-Trinken“) von den bauchseitig gelegenen Geißelzellen aufgenommen.
Auch über das obere Epitheloid (also die „Rückenseite“ des Tieres) können Nahrungspartikel, z. B. ganze Einzeller, aufgenommen werden. Diese Ernährungsweise dürfte im Tierreich einmalig sein: Die in einer Schleimschicht gesammelten Nahrungspartikel werden von den Faserzellen durch Interzellularspalten (Zellzwischenräume) des Epitheloids hindurchgezogen und dann per Phagocytose („Zell-Essen“) verdaut. Ein solches „Einsammeln“ von Nahrungspartikeln durch ein intaktes Abschlussgewebe hindurch ist nur deshalb möglich, weil bei den Placozoa verschiedene „abdichtende“ Elemente (nämlich eine Basallamina unter dem Epitheloid und bestimmte Zell-Zell-Verbindungen) nicht vorhanden sind.
Nicht alle Bakterien, die sich im Inneren der Placozoa finden, werden als Nahrung verdaut: Im endoplasmatischen Reticulum, einem Zellorganell des Fasersynzytiums, befinden sich oft Bakterien, die mit Trichoplax adhaerens anscheinend in Symbiose leben.
Auf fester Oberfläche können sich Placozoa auf zwei verschiedene Arten bewegen: Einmal ermöglicht es ihnen ihre begeißelte Kriechsohle, langsam über den Untergrund hinweg zu gleiten, zum anderen können sie wie eine Amöbe ihre Lage durch Wandlung der Körperform verändern. Die Bewegungen sind dabei nicht zentral koordiniert, da keinerlei Muskel- oder Nervengewebe existiert.
Zwischen Körperform und Fortbewegungsgeschwindigkeit konnte eine enge Verbindung nachgewiesen werden, die zusätzlich abhängig vom Nahrungsangebot ist.
Bei niedriger Nährstoffdichte schwankt die Querschnittsfläche geringfügig, aber unregelmäßig, die Fortbewegungsgeschwindigkeit liegt verhältnismäßig konstant bei etwa 15 Mikrometern pro Sekunde.
Ist die Nährstoffdichte dagegen hoch, oszilliert der Querschnitt mit einer stabilen Periode von etwa 8 Minuten, wobei die größte Ausdehnung des Tiers bis zu doppelt so groß sein kann wie die kleinste. Die Fortbewegungsgeschwindigkeit, die durchweg unterhalb von 5 Mikrometern pro Sekunde liegt, schwankt mit derselben Periode. Eine große Geschwindigkeit entspricht dabei immer einer kleinen Querschnittsfläche und umgekehrt.
Die beiden Ausbreitungsweisen lassen sich, da der Übergang nicht flüssig, sondern abrupt vonstattengeht, sehr gut gegeneinander abgrenzen. Vereinfachend lässt sich Trichoplax adhaerens als nicht-lineares dynamisches System fern dem thermodynamischen Gleichgewicht modellieren.
Eine qualitative Erklärung für das Verhalten des Tiers ergibt sich wie folgt:
Bei niedriger Nährstoffdichte behält Trichoplax eine konstante Geschwindigkeit bei, um ohne unnötigen Zeitverlust Nahrungsquellen ausfindig zu machen.
Ist eine solche Quelle durch hohe Nährstoffdichte identifiziert, erhöht das betreffende Individuum in regelmäßigen Abständen seinen Querschnitt und vergrößert damit die Kontaktfläche mit dem Substrat. Auf diese Weise erweitert sich die Fläche, über die Nährstoffe aufgenommen werden können. Gleichzeitig verringert das Tier seine Geschwindigkeit, um das vorhandene Nahrungsangebot auch tatsächlich auszuschöpfen.
Sobald dies annähernd geschehen ist, verkleinert Trichoplax seinen Querschnitt wieder, um sich weiterzubewegen. Weil Nahrungsquellen wie Algenteppiche oft eine größere Ausdehnung haben, ist es für ein betroffenes Tier sinnvoll, nach kurzer Zeit in der Bewegung innezuhalten, um sich erneut abzuflachen und Nährstoffe aufzunehmen. Daher bewegt sich Trichoplax in dieser Phase nur verhältnismäßig langsam voran.
Die konkrete Richtung, in die sich Trichoplax jeweils bewegt, ist zufallsbestimmt: Misst man, wie schnell sich ein einzelnes Tier von einem (willkürlich bestimmten) Ausgangspunkt wegbewegt, so findet man eine lineare Beziehung zwischen der vergangenen Zeit und der mittleren quadratischen Entfernung von Start- und aktuellem Aufenthaltsort. Ein solcher Zusammenhang charakterisiert auch die zufallsbestimmte Brownsche Bewegung, die somit als Modell für die Fortbewegung der Placozoa dienen kann.
Kleine Tiere sind auch in der Lage, aktiv mit Hilfe ihrer Geißeln zu schwimmen. Sobald sie ein mögliches Substrat berühren, kommt es zu einer Dorsoventralreaktion: Die Geißeln der Rückseite schlagen weiter, während die Geißeln der bauchseitigen Zellen ihren Schlagrhythmus einstellen. Gleichzeitig versucht die Bauchseite, Kontakt mit dem Untergrund herzustellen; kleine Zellaus- und -einstülpungen, die Mikrovilli, die sich an der Oberfläche der säuligen Zellen der Bauchseite befinden, tragen durch ihre adhäsive (klebende) Wirkung zur Anheftung an das Substrat bei.
Eine bemerkenswerte Eigenschaft der Placozoa ist, dass sie sich aus kleinsten Zellbeständen regenerieren können. Selbst wenn im Experiment große Teile des Organismus entfernt werden, entwickelt sich aus dem Rest wieder ein vollständiges Tier. Es ist darüber hinaus möglich, Trichoplax adhaerens durch ein Passiersieb zu streichen, so dass die Einzelzellen zwar nicht zerstört, aber weitgehend voneinander getrennt werden. Im Reagenzglas finden sie sich dann wieder zu kompletten Organismen zusammen. Wird diese Prozedur mit mehreren vorher angefärbten Tieren gleichzeitig vollzogen, geschieht das Gleiche. In diesem Fall kann es sogar vorkommen, dass Zellen, die vorher zu einem bestimmten Tier gehörten, plötzlich als Teil eines anderen wieder auftauchen.
Normalerweise pflanzen sich die Placozoa ungeschlechtlich fort. Das Tier schnürt sich dazu in der Mitte durch, so dass zwei etwa gleich große Tochtertiere entstehen, die nach der Fission allerdings noch für eine Weile lose verbunden bleiben. Seltener findet man Knospungsvorgänge: Dabei trennen sich von der Rückenseite kleine, im Wasser schwebende Zellkugeln („Schwärmer“) ab, die alle bekannten Zelltypen in sich vereinigen und in der Folge zu einem eigenen Individuum heranwachsen.
Die geschlechtliche Fortpflanzung wird möglicherweise durch eine zu große Populationsdichte, in einigen Fällen durch hohe Wassertemperaturen, ausgelöst, sie ist bisher rätselhaft und im Detail kaum verstanden. Im Labor bilden dabei ausschließlich degenerierende, durch Wasseraufnahme anschwellende Individuen binnen vier bis sechs Wochen Eizellen aus. Diese werden vermutlich aus Zellen des unteren Epithels gebildet und wachsen ins Innere des Organismus aus. Dabei schwellen sie durch Aufnahme von Nährstoffen und (extern gebildetem) Dotter durch Phagocytose aus dem sich auflösenden mütterlichen Organismus bis zu einer Größe von 70–120 µm an. Die meisten Individuen bilden nur eine einzige Eizelle, selten kommen bis zu drei vor. Gleichzeitig gebildete, sogenannte F-Zellen stellen möglicherweise die männlichen Geschlechtszellen (Spermien) dar[11], doch wurde weder die Funktion bestätigt noch der Befruchtungsvorgang bisher beobachtet; allerdings werden einige für Spermien typische Proteine in den Zellen exprimiert. Nach der (angenommenen) Befruchtung bildet die Eizelle die sogenannte Befruchtungsmembran, eine schützende Hülle. Die Embryonalentwicklung beginnt mit einer vollständigen, äqualen Furchung. Die jungen Embryonen wachsen im mütterlichen Organismus heran, sie werden durch dessen Degeneration und Auflösung freigesetzt.
Trotz erheblicher Anstrengungen ist es bisher nie gelungen, Embryonen jenseits des 128-Zell-Stadiums am Leben zu erhalten. Die weitere Entwicklung ist daher unbekannt. Vermutlich fehlt in der Laborumgebung ein, bisher unverstandener, kritischer Faktor des natürlichen Lebensraums.
Wegen der Möglichkeit, sich unbegrenzt durch ungeschlechtliche Fortpflanzung zu klonen, ist die Lebensspanne der Placozoa potentiell unendlich; in der Praxis werden einige Entwicklungslinien, die auf ein einziges Tier zurückgehen, seit längerem – in einem Fall seit 1969 – ohne Auftreten sexueller Prozesse in Kultur gehalten.


